
Microbialites are organosedimentary structures formed by microbial communities through binding and trapping and/or in situ precipitation. They have a wide array of distinctive morphologies and a geologic record extending back 3.45 billion years. The origin of morphological variability is hotly debated and elucidating the origins of microfabric differences provides insights into ecosystem functioning and biogeochemistry during much of Earth’s history. We have been studying the role of eukaryotes, and particularly the foraminifera, in modifying microbialites fabrics and impacts on modern microbialites. We propose that the microbialite fossil record, and the demise of stromatolites in the Neoproterozoic may reflect the impact of the concurrent radiation of these protists.
While studying the microbialites of Highborne Cay, in the Bahamas, we became intrigued by the carbonate component which is mostly comprised of ooids. These are small, spherical to ellipsoidal grains characterized by concentric layers of calcium carbonate and organic matter. These sand-sized particles are incorporated with the aid of extra-cellular polymeric substances (EPS), into the matrix of laminated stromatolites and clotted thrombolite mats.
Fossil ooids, oolites, are enigmatic components of the carbonate fossil record including those of the Neoproterozoic. Of particular note, are recent observations that oolitic preserve C-isotopic records of Neoproterozoic environmental change and carbon cycle anomalies (Kristin Bergmann, PhD Dissertation, Caltech, 2013).
Based on analyses of clone libraries of small subunit ribosomal RNA gene fragments and lipid biomarkers, we found that the microbial community colonizing ooid sands was virtually indistinguishable from the community in stromatolites themselves. We also found that ooids from Cat and Andros islands in the Bahamas and from Carbla Beach in Hamelin Pool, Western Australia, preserve abundant and notably similar distributions of hydrocarbons, fatty acids (FAs) and alcohols.
 Histograms showing relative abundances of FAME derived from TLE and TLE2 for three samples of Modern ooids from Joulters Cays and Cat Island in the Bahamas and from North Carbla in Hamelin Pool, Western Australia.
Histograms showing relative abundances of FAME derived from TLE and TLE2 for three samples of Modern ooids from Joulters Cays and Cat Island in the Bahamas and from North Carbla in Hamelin Pool, Western Australia.
Is an ooid a microbialite? We are addressing this question with further molecular studies of ooids from Hamelin Pool and the Bahamian Archipelago.
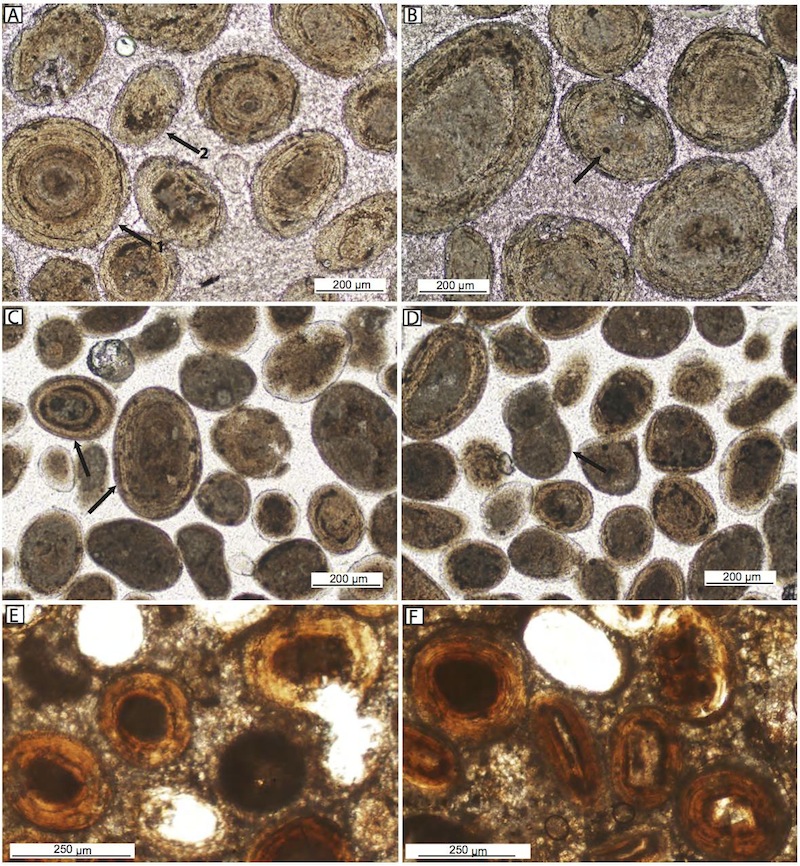
Photomicrograph of ooids embedded in epoxy from Joulter’s Cays and Cat Island, Bahamas, and Carbla Beach, Shark Bay, Australia. A) Joulter’s Cays ooids. Arrow 1 shows ooids with well-preserved outer cortex, and Arrow 2 shows micritized ooid; B) Joulter’s Cays ooids, one with round endolithic boring (arrow); C) Cat Island ooids, with well preserved cortical fabrics (arrows); D) Cat Island ooids, two of which show micritic envelope around ooids (arrow) resulting in grapestone formation; E) and F) Carbla ooids showing well-preserved cortical fabrics and little endolithic boring and/or micritization of laminae.
Collaborators
Our collaborators in the study of microbialites include Linda Jahnke (NASA Ames), Ginny Edgcomb, Joan Bernhard and Mark Roberts (WHOI), Pieter Visscher (UConn), Sara Pruss (Smith), Tanja Bosak (MIT) and Steven Beaupre (Harvard).
Funding sources
This research has been supported by the NSF Chemical Oceanography Program (OCE-0926372) and the NASA Astrobiology Institute (NNA08CN84A and NNA13AA90A).
References
- Edgcomb, V.P., Bernhard J.M., Beaudoin D., Pruss S., Welander P.V., Schubotz F., Mehay S., Gillespie A.L. and Summons R.E. (2013) Molecular indicators of microbial diversity in oolitic sands of Highborne Cay, Bahamas. Geobiology 11, 234–251.
- Bernhard J.M., Edgcomb V.P., McIntyre-Wressnig A., Summons R.E. , Visscher P.T., Jeglinski, M., 2013. Insights into foraminiferal influences on microfabrics of microbialites at Highborne Cay, Bahamas. Proceedings of the National Academy of Sciences (USA) 110, 9830-9834.
- Summons R.E., Bird L.R., Gillespie A.L., Pruss S.B., Roberts M. and Sessions A.L., 2013. Lipid biomarkers in ooids from different locations and ages: evidence for a common bacterial flora Geobiology 11, 420–436.
- Edgcomb V.P., Bernhard J.M., Summons R.E., Orsi W., Beaudoin D. and Visscher P.T., 2013. Active eukaryotes in microbialites from Highborne Cay, Bahamas and Hamelin Pool (Shark Bay), Australia. ISME J doi:10.1038/ismej.2013.130.
- Schubotz F., Meyer-Dombard D.R., Bradley A.S., Fredricks H.F., Shock E.L., Hinrichs K.-U., Summons R.E., 2013. The Lipid and Isotopic Compositions of Streamer Biofilm Communities in the Lower Geyser Basin of Yellowstone National Park. Geobiology, 11, 549-569.
- Jahnke L.L, Turk-Kubo K.A., Parenteau M.N., Green S.J., Kubo M.D.Y, Vogel M., Summons R.E. and Des Marais D.J., 2013. Molecular and Lipid Biomarker Analysis of a Gypsum-hosted Endoevaporitic Microbial Community. Geobiology 12, 62-82.
